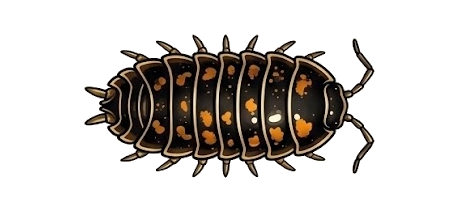
Order · Diplopoda · Pentazonia · Oniscomorpha

Order
Glomerida
Northern pill millipedes — armed with the most potent alkaloids in Diplopoda
0Described Species
0Genera
0Body Tergites
0Year Max Lifespan
0Months to Replenish Defense
Scroll